Recombinant adeno-associated virus (AAV) vector has been increasingly used as a vehicle in gene therapy to deliver therapeutic genes.1 During the AAV biomanufacturing process, high percentage of capsids that do not contain the desired DNA (empty capsids) are also produced and are therefore unable to provide therapeutic benefits. It has been reported that empty capsids can reduce transduction efficiency and induce unnecessary immune response.2 On the other hand, recent data show that empty AAV capsids can enhance gene transfer.3 Whether beneficial or detrimental, full versus empty capsid formulations need to be monitored to ensure consistent product quality and efficacy.
Multiple techniques have been used to quantify empty capsids in AAV samples. Analytical ultracentrifugation (AUC),4 UV absorbance spectrophotometry,5 and electron microscopy6,7 are extensively used in measuring empty/full ratio of AAV particles. AUC, which measures the sedimentation rate of the particles, is the gold standard in this application area. However, hours are required for skilled AUC proficient analysts to interpret the data and AUC deployment can consume hundreds of microliters of highly valued sample. In addition, an assay based on AUC analysis is challenging to validate. Static spectrophotometry that measures at 260 nm versus 280 nm has also been used to estimate empty from full capsids. Although a quicker method, the accuracy of the results can be compromised by sample matrix interferences. In addition, sizeable amount of sample is required to obtain adequate absorbance values. Transmission electron microscopy (TEM) and cryogenic electron microscopy (Cryo-EM) provide direct visualization and counting of the empty and full particles, but the quantification heavily relies on image quality and field selection. The most recently used technique for this analysis is charge detection mass spectrometry (CDMS).8 This MS method directly measures the molecular weight of the particles and has been shown capable of accurately measuring empty, partial, and full AAV particles. Currently, no commercial CDMS instrument is available.
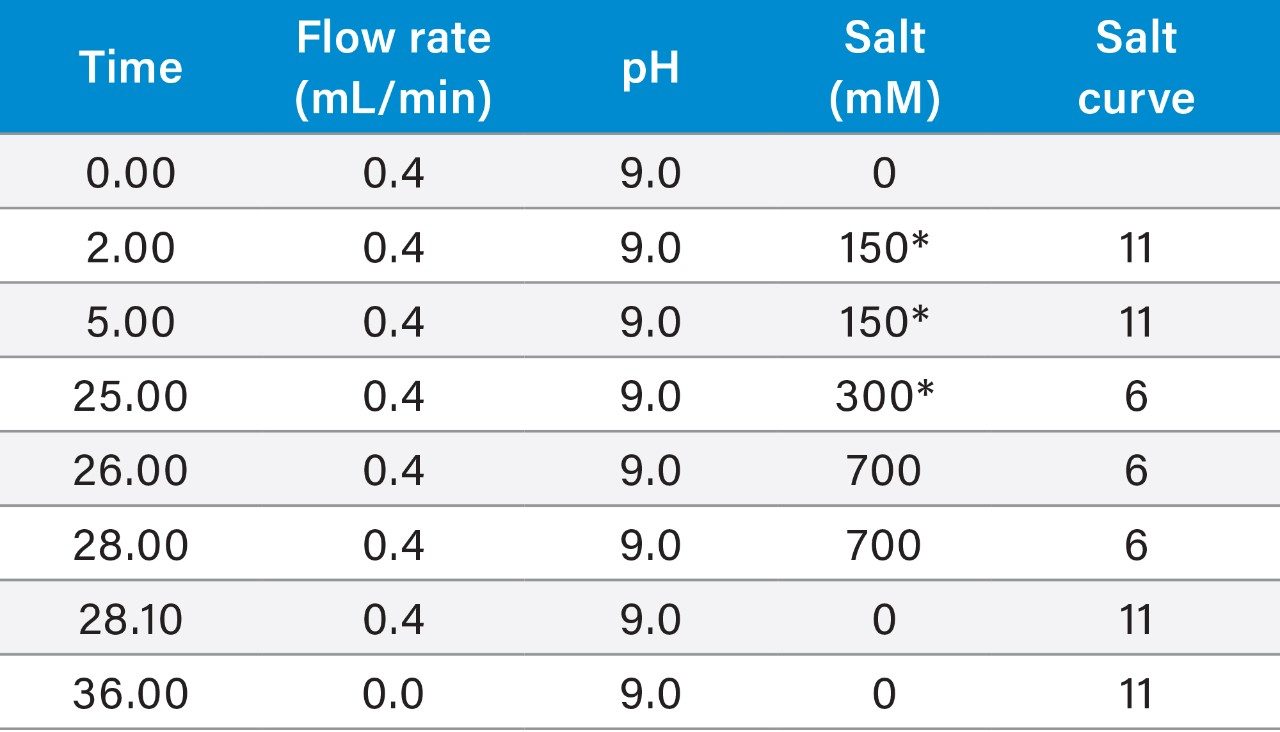
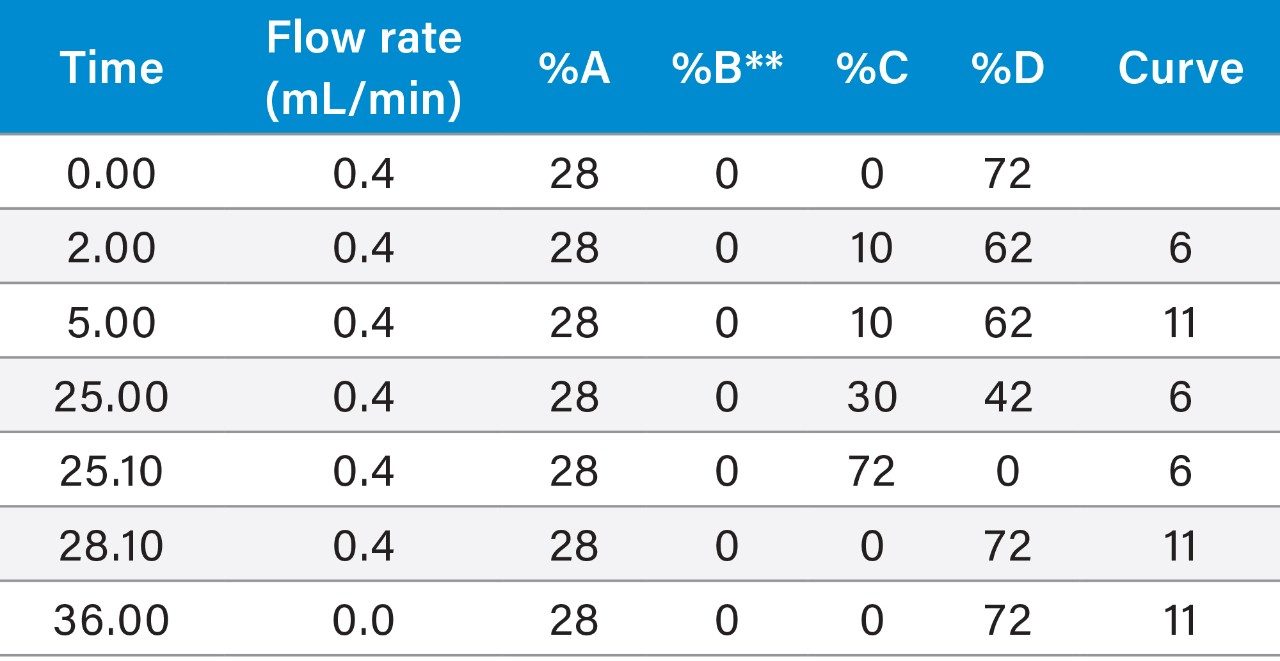
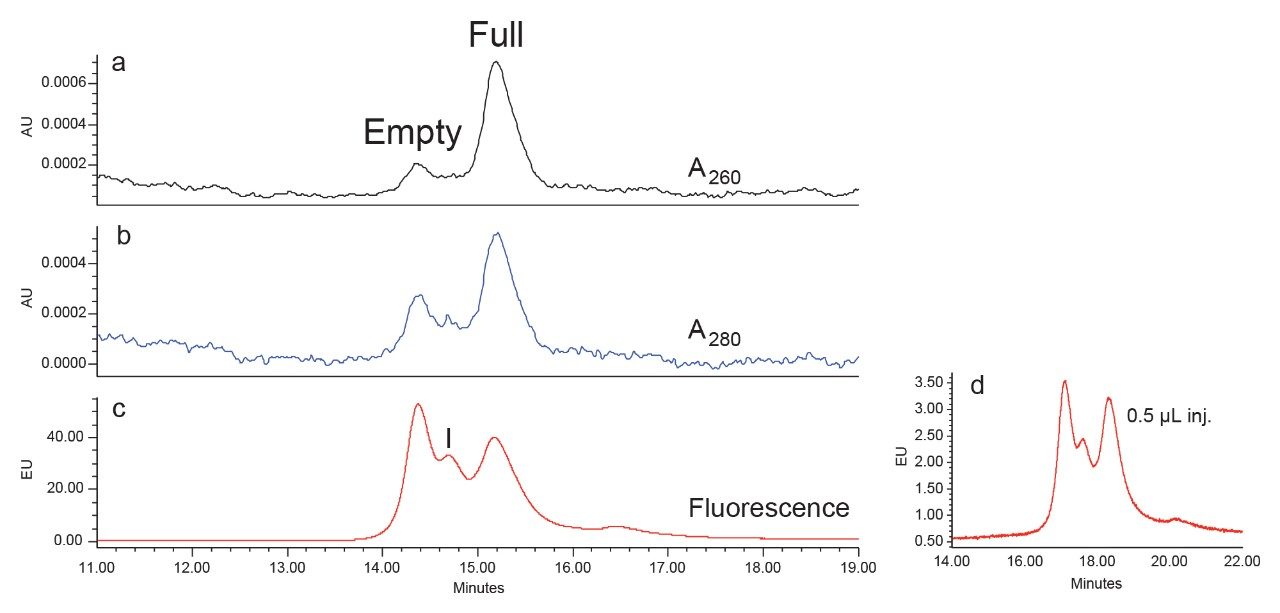
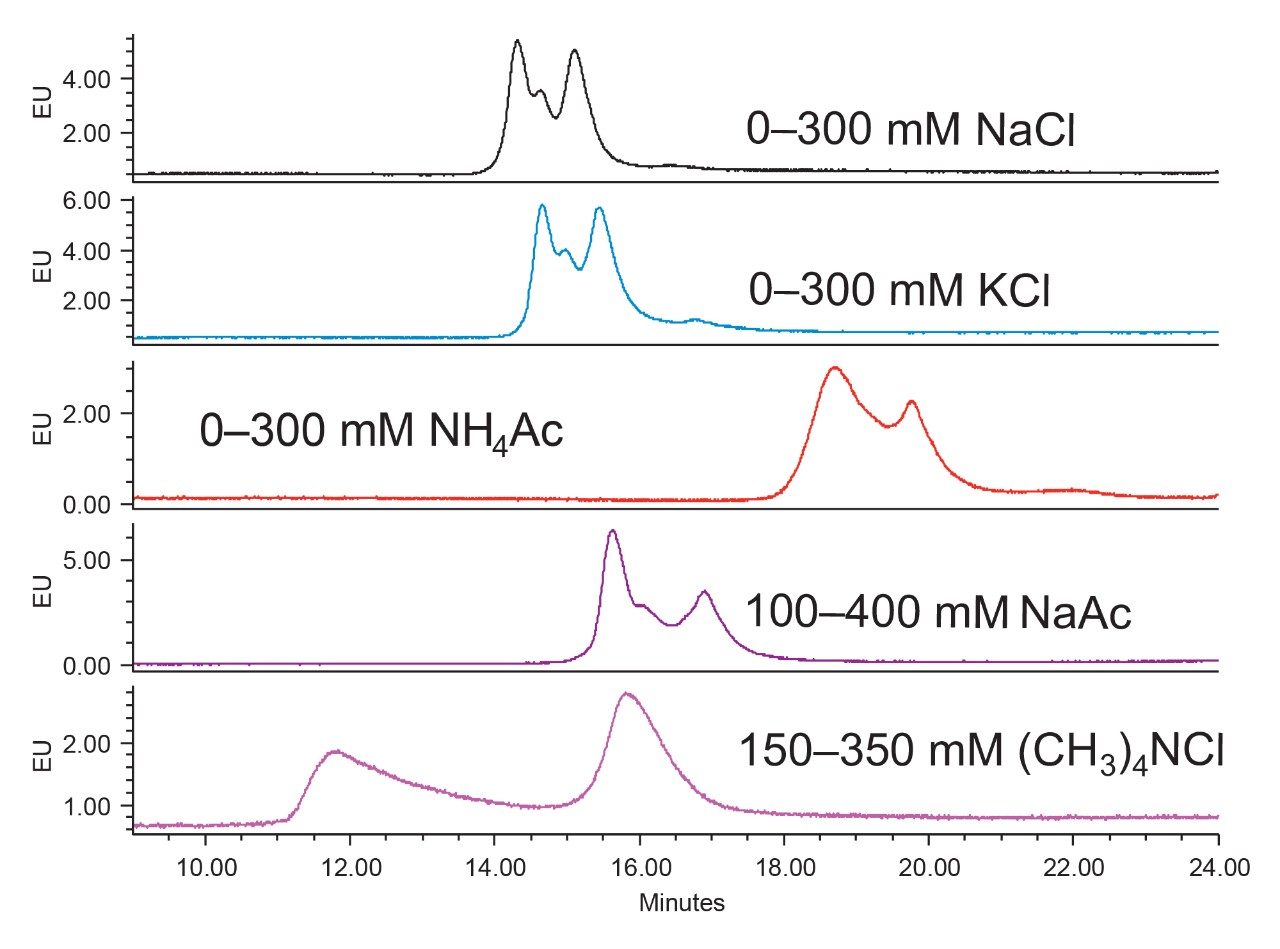
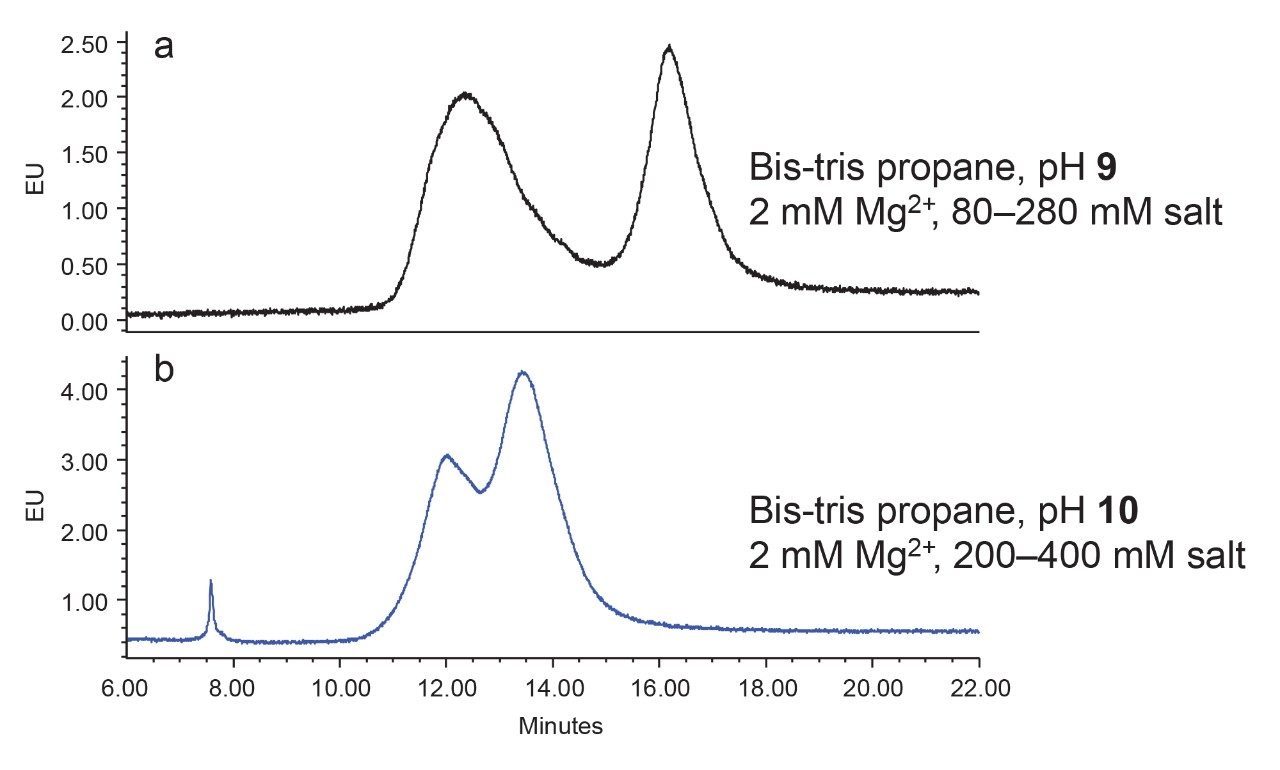
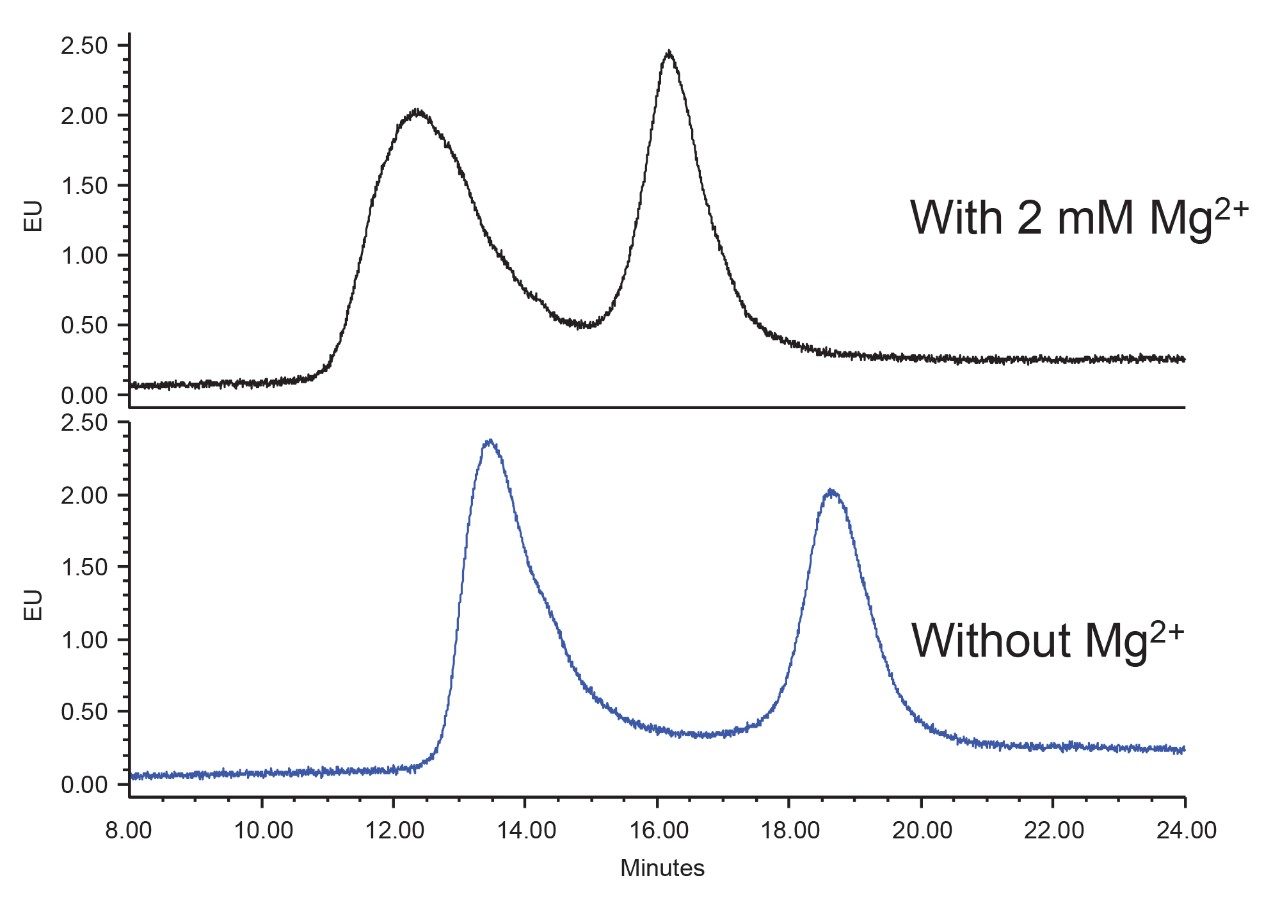
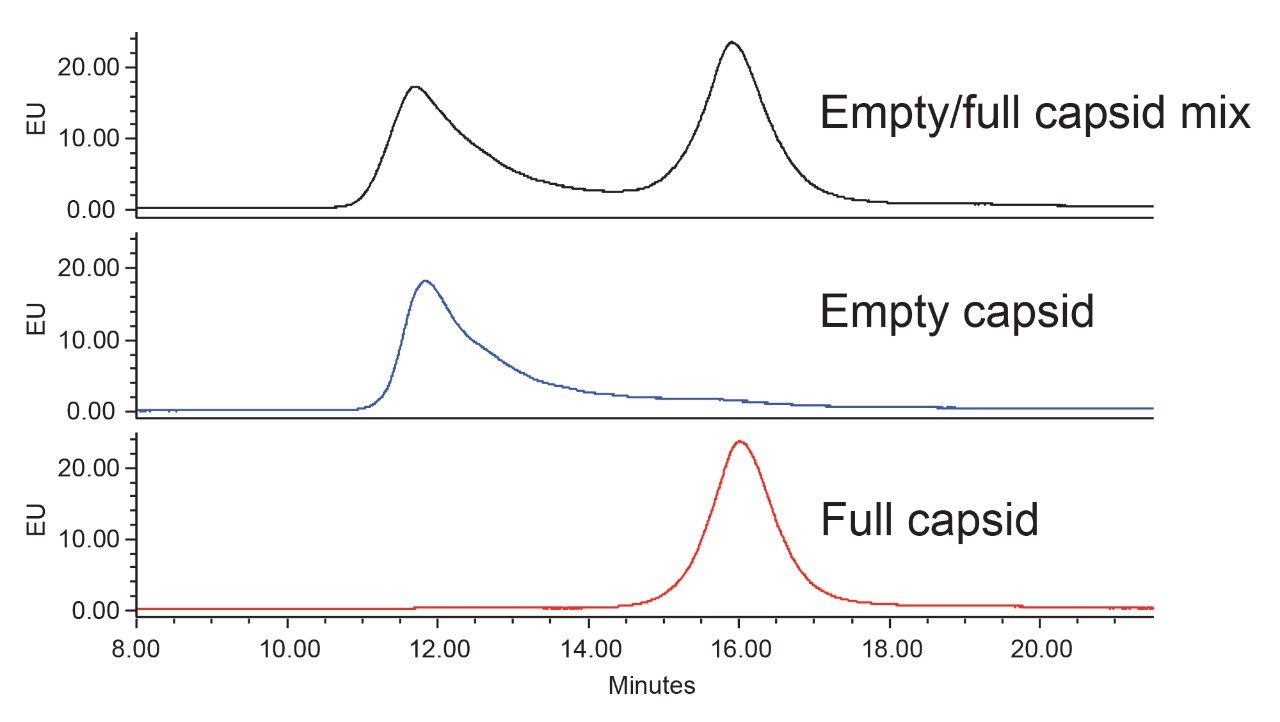
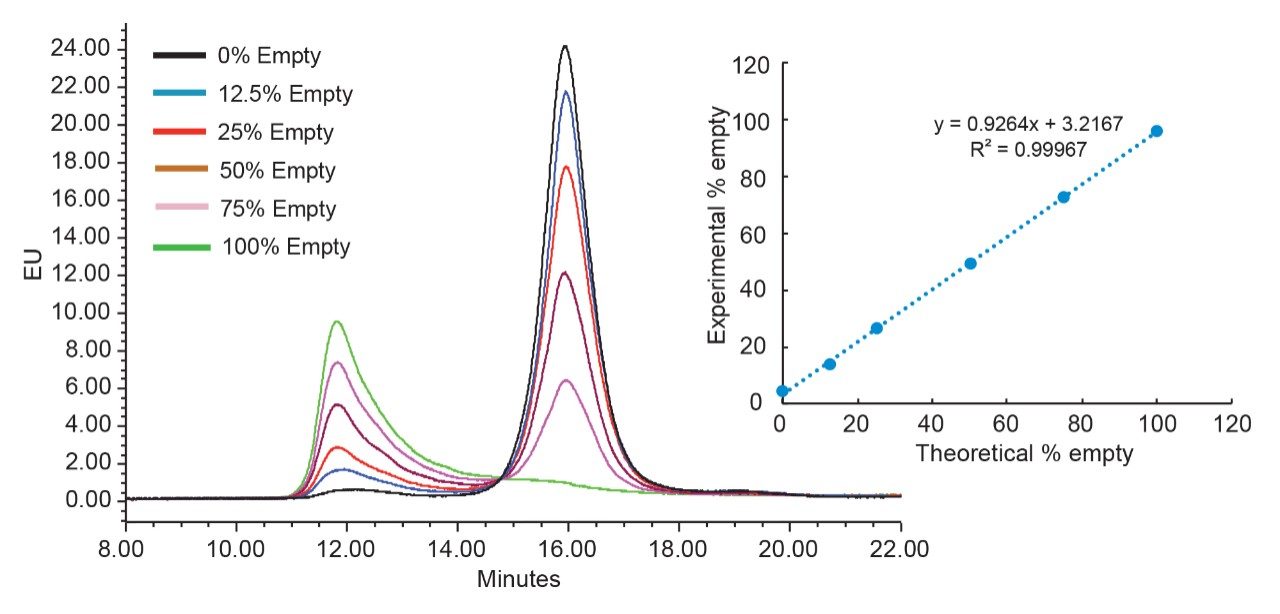
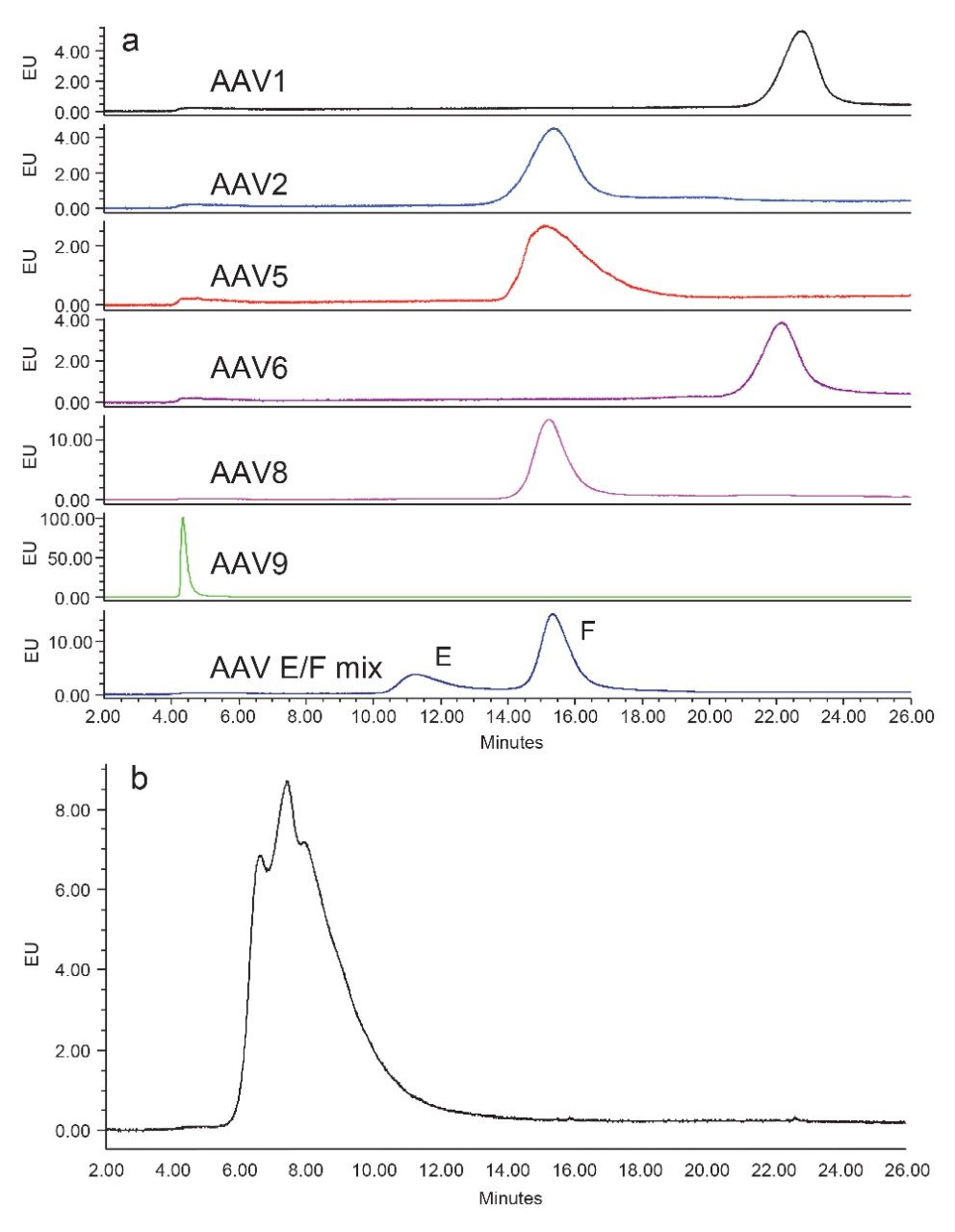
Anion-exchange chromatography (AEX) was reported to have the potential of separating empty and full capsids and it has been suggested that this separation is driven by differences in their relative surface charges.9,10 This technique consumes small amounts of sample, and can yield robust and reproducible results using standard LC instrumentation. In this application note, we show the separation and quantification of empty and full capsids on a Waters Protein-Pak Hi Res Q AEX Column.